























Les grands dauphins sont probablement les espèces de dauphins les plus connues, celles sur lesquelles repose l’image classique que chacun se fait d’un « dauphin ». L’emblématique Flipper, star des films et émissions de télévision depuis les années 1960, était un grand dauphin, comme la majorité des dauphins captifs dans les delphinariums du monde entier. On trouve des grands dauphins dans presque tous les océans et toutes les mers, à l’exception des eaux les plus froides des pôles. Deux espèces de grands dauphins sont actuellement reconnues, et dans certains cas, elles peuvent être présentes dans la même région. Il s’agit du grand dauphin commun (Tursiops truncatus) et du grand dauphin de l’Indo-Pacifique (Tursiops aduncus). En plus de cette distinction entre espèces (voir plus de détails ci-dessous), il existe une grande variation entre les populations côtières et hauturières du grand dauphin commun dans les différentes régions du monde. Les populations diffèrent fortement par la longueur moyenne, le poids, les nageoires, la coloration, le régime alimentaire et le comportement. Une troisième espèce possible – le dauphin de Burrunan – a été proposée pour les populations côtières de grands dauphins en Australie1, et les scientifiques et généticiens du monde entier consacrent beaucoup d’efforts à mieux comprendre la taxonomie des grands dauphins. Comme les humains, les différentes populations vivent dans des environnements très divers et ont adapté leur régime alimentaire, leurs comportements et leurs caractéristiques externes à ces habitats2. Cependant, où qu’ils se trouvent, les grands dauphins ont tendance à être parmi les espèces de dauphins les plus accessibles et les plus actives, ce qui en fait une cible populaire pour les activités d’observation des dauphins.
Partager cette page!
XPartager cette page sur les médias sociaux:
Grand dauphin Tursiops truncatus et T. aduncus

Risques de confusion
De manière quelque peu déroutante, les grands dauphins sont parfois appelés marsouins, notamment dans le sud-est des États-Unis. Cependant, les véritables espèces de marsouins sont toutes beaucoup plus petites et se caractérisent par un museau émoussé (sans bec) et des dents en forme de pique. En raison de leur présence cosmopolite, l’aire de répartition des grands dauphins chevauche celle de nombreuses autres espèces de dauphins. Dans l’Indo-Pacifique tropical et le long de la côte ouest de l’Afrique, la répartition des populations côtières de grands dauphins peut recouper celle des dauphins à bosse. Les dauphins à bosse se distinguent des grands dauphins par la large bosse sous leur aileron dorsal généralement plus court et moins incurvé que celui des grands dauphins, et par leur bec plus long et plus fin. Les grands dauphins peuvent aussi être confondus avec le sténo rostré, de taille et de silhouette similaire, mais dont le front est en pente douce sans pli prononcé entre le rostre et le front. Les dauphins tachetés pantropicaux ou de l’Atlantique ne sont pas toujours très tachetés et peuvent alors être confondus avec les grands dauphins, mais ils ont généralement un bec plus long et plus fin.
Répartition
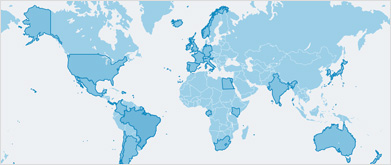
Le grand dauphin commun vit dans presque toutes les régions tropicales et tempérées, aussi bien dans les eaux côtières qu’en haute mer3. Il est présent dans la plupart des mers fermées ou semi-fermées (p. ex. la mer du Nord, la Méditerranée, la mer Noire, le golfe Persique), et il fréquente les baies, les lagunes, les chenaux ainsi que les embouchures des cours d’eau4. Le grand dauphin de l’Indo-Pacifique a une aire de répartition plus restreinte dont les limites se situent à l’ouest à l’extrémité sud de l’Afrique, et à l’est aux îles Salomon/Nouvelle-Calédonie. Sa présence est généralement limitée aux eaux côtières du plateau continental, bien qu’on le rencontre également autour de certains groupes d’îles de l’Indo-Pacifique4.
Le grand dauphin commun est présent dans les pays et territoires suivants : Afrique du Sud ; Albanie ; Algérie ; Allemagne ; Angola ; Anguilla ; Antigua-et-Barbuda ; Arabie saoudite ; Argentine ; Aruba ; Australie ; Bahamas ; Bahreïn ; Bangladesh ; Barbade ; Belgique ; Belize ; Bénin ; Bermudes ; Bonaire, Saint-Eustache et Saba (Saba, Saint-Eustache) ; Bosnie-Herzégovine ; Brésil ; Brunéi Darussalam ; Bulgarie ; Cabo Verde ; Cambodge ; Cameroun ; Canada ; Chili ; Chine ; Chypre ; Colombie ; Comores ; Corée (République de) ; Costa Rica ; Côte d’Ivoire ; Croatie ; Cuba ; Curaçao ; Danemark ; Djibouti ; Dominique ; El Salvador ; Émirats arabes unis ; Équateur ; Égypte ; Espagne ; États-Unis d’Amérique ; Fédération de Russie ; Fidji ; France ; Gabon ; Gambie ; Géorgie ; Ghana ; Gibraltar ; Grèce ; Grenade ; Guadeloupe ; Guam ; Guatemala ; Guernesey ; Guinée ; Guinée-Bissau ; Guyana ; Guyane française ; Haïti ; Honduras ; Hong Kong ; Île de Man ; Îles Caïmans ; Îles Cocos (Keeling) ; Îles Cook ; Îles Féroé ; Îles Malouines (Falkland/Malvinas) ; Îles Mariannes septentrionales ; Îles Marshall ; Îles Pitcairn ; Îles Salomon ; Îles Turques et Caïques ; Inde ; Indonésie ; Iran (République islamique d’) ; Irlande ; Israël ; Italie ; Jamaïque ; Japon ; Jersey ; Kenya ; Kiribati ; Koweït ; Liban ; Liberia ; Libye ; Madagascar ; Malaisie ; Maldives ; Malte ; Maroc ; Martinique ; Mauritanie ; Mayotte ; Mexique ; Micronésie (États fédérés de) ; Monaco ; Monténégro ; Mozambique ; Myanmar ; Namibie ; Nauru ; Nicaragua ; Nigéria ; Nioué ; Nouvelle-Calédonie ; Nouvelle-Zélande ; Oman ; Pakistan ; Palaos ; Panama ; Papouasie–Nouvelle-Guinée ; Pays-Bas ; Pérou ; Philippines ; Polynésie française ; Porto Rico ; Portugal ; Qatar ; République arabe syrienne ; République dominicaine ; Réunion ; Roumanie ; Royaume-Uni de Grande-Bretagne et d’Irlande du Nord ; Sahara occidental ; Sainte-Hélène, Ascension et Tristan da Cunha ; Sainte-Lucie ; Sainte-Lucie ; Saint-Kitts-et-Nevis ; Saint-Martin (partie française) ; Saint-Martin (partie néerlandaise, Sint Maarten) ; Saint-Pierre-et-Miquelon ; Saint-Vincent-et-les-Grenadines ; Samoa ; Samoa américaines ; Sao Tomé-et-Principe ; Sénégal ; Seychelles ; Singapour ; Slovénie ; Somalie ; Sri Lanka ; Suriname ; Taiwan (Province de Chine) ; Tanzanie (République-Unie de) ; Thaïlande ; Togo ; Tonga ; Trinité-et-Tobago ; Tunisie ; Turquie ; Ukraine ; Uruguay ; Vanuatu ; Venezuela (République bolivarienne du) ; Viet Nam ; Wallis et Futuna ; Yémen.
Le grand dauphin de l’Indo-Pacifique est présent dans les pays et territoires suivants : Afrique du Sud ; Arabie saoudite ; Australie ; Bahreïn ; Bangladesh ; Brunéi Darussalam ; Cambodge ; Chine ; Comores ; Égypte ; Émirats arabes unis ; Érythrée ; Îles Salomon ; Inde ; Indonésie ; Iran (République islamique d’) ; Japon ; Kenya ; Madagascar ; Malaisie ; Mayotte ; Mozambique ; Myanmar ; Oman ; Pakistan ; Papouasie–Nouvelle-Guinée ; Philippines ; Singapour ; Somalie ; Sri Lanka ; Taiwan (Province de Chine) ; Tanzanie (République-Unie de) ; Thaïlande ; Timor-Leste ; Yémen.
Biologie et écologie
Alimentation
Les grands dauphins se nourrissent d’une grande variété de proies, les différentes populations privilégiant diverses espèces de poissons, de calmars et parfois de crustacés en fonction de l’habitat qu’elles occupent3,4. Les populations proches du rivage préfèrent généralement les poissons de fond et les poissons produisant des sons comme certains sciaenidés et les gorettes3,5,6, tandis que d’autres populations (y compris en haute mer) consomment des poissons vivant en bancs comme les mulets et d’autres espèces, dont les maquereaux, les thons et les bonites 3,7. Les grands dauphins emploient toute une gamme de techniques de chasse pouvant être hautement spécialisées et créatives, comme l’utilisation d’éponges pour protéger leur bec lorsqu’ils creusent dans le sable à la recherche de proies8, ainsi que le rabattage des poissons sur la plage (strand feeding en anglais), une chasse coopérative au cours de laquelle les dauphins nagent ensemble en ligne pour repousser les poissons vers le rivage, s’échouent partiellement pour capturer leurs proies qui s’agitent sur la grève, puis glissent de nouveau dans l’eau.
Structure sociale, reproduction et croissance
Les grands dauphins peuvent se rencontrer en petits groupes de 2 à 15 individus, ou en groupes pouvant compter jusqu’à 1000 individus, les populations côtières présentant généralement des groupes plus réduits que les populations hauturières3. Dans de nombreuses baies, détroits et estuaires où ils ont été étudiés, comme dans la baie de Sarasota en Floride, il a été constaté que les grands dauphins communs étaient présents toute l’année, restant dans la même zone pendant plusieurs décennies et de nombreuses générations9. Ils sont très loquaces et produisent trois grandes catégories de sons : des sifflements, des clics d’écholocalisation et des sons pulsés en rafale. Leurs vocalisations ont été étudiées tant chez des individus captifs que chez des individus sauvages. Elles comprennent un « sifflement signature », unique et propre à chaque individu, qui semble être utilisé pour communiquer leur identité, leur localisation et peut-être même leur état émotionnel10.
Chez le grand dauphin commun, la femelle atteint la maturité sexuelle entre 5 et 13 ans, le mâle entre 9 et 14 ans3. Le petit naît environ 12,5 mois après la fécondation et se nourrit du lait de sa mère pendant les deux premières années de sa vie. Il commence à compléter son alimentation par des aliments solides à partir de 4 à 6 mois et sa mère lui enseigne activement comment chasser. La femelle s’occupe de son jeune pendant 3 à 6 ans, leur séparation ayant généralement lieu à la naissance du petit suivant3.
Il a été montré que des hiérarchies existent entre les mâles et que cela entraîne des interactions agressives. Toutefois, les grands dauphins de l’Indo-Pacifique mâles peuvent aussi coopérer entre eux pour contraindre une femelle à s’accoupler11 et les grands dauphins communs mâles agissent souvent au sein d’une alliance forte et à long terme pour protéger les femelles réceptives des approches d’autres mâles12. Les mâles portent généralement plus de marques que les femelles et présentent davantage de cicatrices sur leur aileron dorsal, y compris des marques de dents caractéristiques provenant d’interactions avec d’autres dauphins mâles13.
Recherche, menaces et conservation
Une grande partie de ce que nous savons sur la biologie, les capacités d’apprentissage et la structure sociale des dauphins a été apprise en étudiant de près les nombreux grands dauphins maintenus en captivité dans le monde entier. Ces dauphins ont été utilisés dans des contextes expérimentaux pour comprendre comment fonctionne leur écholocalisation, comment ils communiquent entre eux, comment ils « dorment » et comment ils acquièrent de nouvelles compétences. Ils sont également utilisés par l’armée pour effectuer des tâches sous-marines considérées comme trop risquées pour les plongeurs humains, et sont les espèces les plus souvent entraînées dans les delphinariums du monde entier.
Cependant, les chercheurs disposent également de techniques de plus en plus sophistiquées pour étudier les dauphins dans leur milieu naturel. Les études par photo-identification, dans lesquelles des dauphins sont photographiés et reconnus individuellement grâce aux entailles et cicatrices uniques marquant leur aileron dorsal, permettent aux chercheurs de suivre les déplacements et les histoires de vie des individus au cours du temps. Dans les populations bien étudiées, comme celles des grands dauphins de la baie de Sarasota en Floride, les individus et leurs communautés ont été suivis pendant plus de 48 ans, ce qui a permis aux chercheurs de construire des arbres généalogiques et de mesurer les paramètres du cycle de vie, le succès de reproduction ainsi que les taux de natalité et de mortalité9. L’utilisation d’hydrophones pour l’enregistrement des vocalisations, l’échantillonnage génétique, le marquage pour le suivi satellitaire et de nombreuses autres techniques permettent d’acquérir des connaissances supplémentaires précieuses sur les populations et sur la manière de les protéger. Pour en savoir plus sur ces techniques, cliquer ici.
Prédateurs naturels
Les principaux prédateurs naturels des grands dauphins sont les requins, et des études par photo-identification ont révélé des cicatrices associées à des attaques de requins dans plusieurs populations14-16. Les piqûres de raies et les agressions entre dauphins ont également été signalées comme causes de mortalité dans certains cas3.
Menaces d’origine humaine
Comme pour toutes les baleines et tous les dauphins, les prises accessoires, c’est-à-dire l’enchevêtrement accidentel dans les engins de pêche, constituent la principale source de mortalité d’origine humaine pour les grands dauphins. Cela est particulièrement important dans les zones côtières où les filets maillants à grandes mailles sont les principaux engins de pêche utilisés, car ils sont souvent posés et laissés sans surveillance pendant de longues périodes, provoquant l’enchevêtrement des dauphins lorsqu’ils se déplacent ou lorsqu’ils chassent leurs proies dans les filets17-20. Dans certaines zones côtières, les engins de pêche de loisir sont la principale source de mortalité et de blessures graves21. De plus, les ruissellements et les écoulements agricoles et industriels dans les zones d’habitation humaine dense entraînent des niveaux élevés de contaminants dans les populations côtières de grands dauphins 22-25, et l’augmentation des charges en contaminants a été reliée à une mortalité élevée des nouveau-nés en raison des fortes concentrations de ces substances dans le lait maternel26. La mortalité de grands dauphins a également été reliée à des proliférations d’algues toxiques, parfois appelées « marées rouges », qui peuvent se produire naturellement, mais sont parfois provoquées par des ruissellements et des rejets provenant de zones fortement peuplées ou cultivées27,28.
État de conservation
Il est difficile d’attribuer un état de conservation aux grands dauphins communs et de l’Indo-Pacifique à l’échelle de la planète ou de l’aire de répartition. Le grand dauphin de l’Indo-Pacifique est particulièrement difficile à évaluer, car il a tendance à constituer des populations côtières fragmentées, et son aire de répartition comprend de nombreux pays où il y a eu peu ou pas de recherches formelles. À ce titre, il est classé dans la catégorie Données insuffisantes sur la Liste rouge des espèces menacées de l’UICN. Le grand dauphin commun est classé par l’UICN au niveau mondial dans la catégorie Préoccupation mineure, mais cela peut être trompeur, car de nombreuses populations subissent de graves déclins, notamment : la population de grands dauphins de la Méditerranée considérée comme Vulnérable à l’extinction ; et celle de grands dauphins de la mer Noire, classée dans la catégorie En danger et inscrite à l’Annexe I de la CMS en raison de la chasse, des captures d’individus vivants et des taux élevés de prises accessoires par les pêcheries. La population de grands dauphins du Fjordland, qui comprend Doubtful Sound en Nouvelle-Zélande, est considérée comme En danger critique d’extinction 29,30, et il est possible que ce déclin soit lié à la pression exercée par les activités d’observation des dauphins31. En général, les populations côtières et sédentaires de grands dauphins sont les plus exposées au risque de déclin de leur santé ou de leurs effectifs en raison du chevauchement de leur habitat avec diverses activités humaines, notamment la pêche, l’agriculture, l’industrie et le trafic maritime.
Grands dauphins et tourisme d’observation
Parce qu’on les trouve très souvent là où les gens vont sur l’eau, et parce qu’ils ont tendance à être accessibles, curieux et intéressants à observer, les grands dauphins sont souvent la cible d’activités d’observation. Les cas de grands dauphins solitaires qui commencent à interagir fréquemment avec les humains et les rencontres qui ont lieu dans le cadre de programmes de nourrissage de dauphins captifs, semi-captifs ou sauvages renforcent également l’idée que cette espèce recherche et apprécie la compagnie humaine. Il est très préoccupant que les petites populations côtières de grands dauphins qui sont présentes toute l’année et disposent d’une zone relativement restreinte où se nourrir, se reposer et socialiser puissent être gravement affectées par des activités irresponsables d’observation des dauphins. Les populations sédentaires de grands dauphins, comme celles de Bocas del Toro, au Panama, peuvent être exposées quotidiennement pendant des heures à la présence continue de bateaux et au bruit des moteurs, et perdre régulièrement des occasions importantes de se nourrir ou de se reposer, ce qui nuit à leur santé générale 32,33. Certaines études ont établi un lien entre la pression intense exercée par le tourisme d’observation et le déclin ou le départ des grands dauphins de la zone où ils étaient observés34,35. Ainsi, il est extrêmement important que les activités d’observation des dauphins n’aggravent pas les menaces, mais contribuent au contraire à leur conservation en respectant des lignes directrices d’observation responsable des espèces.
Références
Afficher / Masquer les références
- Charlton-Robb, K. et al. A New Dolphin Species, the Burrunan Dolphin Tursiops australis sp. nov., Endemic to Southern Australian Coastal Waters. PLoS ONE 6, e24047, doi:10.1371/journal.pone.0024047 (2011).
- IWC. Report of the Scientific Committee: Annex M: Report of the Sub-Committee on Small Cetaceans. 40 (International Whaling Commission, Bled, Slovenia, 2017).
- Wells, R. S. & Scott, M. D. Common bottlenose dolphin, Tursiops truncatus, in Encyclopedia of Marine Mammals (eds W. Perrin, B. Wursig, & J.G.M. Thewissen) 249-255 (Elsevier, 2009).
- Jefferson, T. A., Webber, M. A. & Pitman, R. L. Marine Mammals of the World: a Comprehensive Guide to their Identification. Second Edition. (San Diego: Academic Press, 2015).
- Ponnampalam, L. et al. Stomach contents of small cetaceans stranded along the Sea of Oman and Arabian Sea coasts of the Sultanate of Oman. Journal of the Marine Biological Association of the UK 92, 1699-1710 (2012).
- Ross, G. B. G. The smaller cetaceans of the south east coast of Southern Africa. Ann. of the Cape Province Museum (Natural History) 15, 173-410 (1984).
- Wang, J., Y. & Yang, S. C. Indo-Pacific Bottlenose dolphin, Tursiops aduncus, in Encyclopedia of Marine Mammals (ed W. F. Perrin) 602-608 (2009).
- Krützen, M. et al. Cultural transmission of tool use by Indo-Pacific bottlenose dolphins (Tursiops sp.) provides access to a novel foraging niche. Proceedings of the Royal Society B: Biological Sciences 281, doi:10.1098/rspb.2014.0374 (2014).
- Wells, R. S. Social Structure and Life History of Bottlenose Dolphins Near Sarasota Bay, Florida: Insights from Four Decades and Five Generations, in Primates and Cetaceans: Field Research and Conservation of Complex Mammalian Societies (eds Juichi Yamagiwa & Leszek Karczmarski) 149-172 (Springer Japan, 2014).
- Janik, V. M., Sayigh, L. S. & Wells, R. S. Signature whistle shape conveys identity information to bottlenose dolphins. PNAS 103, 8293-8297 (2006).
- Scott, E. M., Mann, J., Watson-Capps, J. J., Sargeant, B. L. & Connor, R. C. Aggression in bottlenose dolphins: evidence for sexual coercion, male-male competition, and female tolerance through analysis of tooth-rake marks and behaviour. Behaviour 142, 21-44 (2005).
- Wells, R. S. Dolphin social complexity: Lessons from long-term study and life history, in Animal Social Complexity: Intelligence, Culture, and Individualized Societies (eds F.B.M. de Waal & P.L. Tyack) 32-56 (Harvard University Press, 2003).
- Marley, S. A., Cheney, B. & Thompson, P. M. Using tooth rakes to monitor population and sex differences in aggressive behaviour in bottlenose dolphins (Tursiops truncatus). Aquatic mammals 39, 107 %@ 0167-5427.
- Corkeron, P. J., Morris, J. G. & Bryden, H. L. Interactions between bottlenose dolphins and sharks in Moreton Bay, Queensland. Aquatic Mammals 13, 109-113 (1987).
- Heithaus, M. R. et al. Spatial variation in shark-inflicted injuries to Indo-Pacific bottlenose dolphins (Tursiops aduncus) of the southwestern Indian Ocean. Marine Mammal Science 33, 335-341, doi:10.1111/mms.12346 (2017).
- Smith, F., Allen, S. J., Bejder, L. & Brown, A. M. Shark bite injuries on three inshore dolphin species in tropical northwestern Australia. Marine Mammal Science, n/a-n/a, doi:10.1111/mms.12435 (2017).
- Amir, O. A., Berggren, P. & Jiddawi, N. S. The incidental catch of dolphins in gillnet fisheries in Zanzibar, Tanzania. Western Indian Ocean Journal of Marine Science 1, 155-162 (2002).
- Friedlaender, A. S., McLellan, W. A. & Pabst, D. A. Characterising an interaction between coastal bottlenose dolphins (Tursiops truncatus) and the spot gillnet fishery in southeastern North Carolina, USA. Journal of Cetacean Research and Management 3, 293-304 (2001).
- Lopez, B. D. Interactions between Mediterranean bottlenose dolphins (Tursiops truncatus) and gillnets off Sardinia, Italy. ICES Journal Marine Science 63, 946-951 (2006).
- Van Waerebeek, K. et al. On the status of the common bottlenose dolphin Tursiops truncatus in western Africa, with emphasis on fisheries interactions, 1947-2015. 20 (2016).
- Wells, R. S. et al. Consequences of injuries on survival and reproduction of common bottlenose dolphins (Tursiops truncatus) along the west coast of Florida. Marine Mammal Science 24, 774-794, doi:10.1111/j.1748-7692.2008.00212.x (2008).
- Adams, J. et al. Land use and the spatial distribution of perfluoroalkyl compounds as measured in the plasma of bottlenose dolphins (Tursiops truncatus). Marine Environmental Research 66, 430–437 (2008).
- Hansen, L. J. et al. Geographic Variation in Polychorinated Biphenyl and Organochlorine Pesticide Concentrations in the Blubber of Bottlenose Dolphins from the U.S. Atlantic Coast. Science of the Total Environment 319, 147-172 (2004).
- Pompe-Gotal, J., Srebocan, E., Gomercic, H. & Prevendar Crnic, A. Mercury concentrations in the tissues of bottlenose dolphins (Tursiops truncatus) and striped dolphins (Stenella coeruloalba) stranded on the Croatian Adriatic coast. Veterinarni Medicina 54, 598-604 (2009).
- Shoham-Frider, E. et al. Persistent organochlorine pollutants and heavy metals in tissues of common bottlenose dolphin (Tursiops truncatus) from the Levantine Basin of the Eastern Mediterranean. Chemosphere 7, 621–627 (2009).
- Wells, R. S. et al. Bottlenose dolphins as marine ecosystem sentinels: Developing a health monitoring system. EcoHealth 1, 246-254 (2004).
- Fire, S. E. et al. Brevetoxin-associated mass mortality event of bottlenose dolphins and manatees along the east coast of Florida, USA. MEPS 526, 241-251 (2015).
- Fire, S. E. et al. Co-occurrence of multiple classes of harmful algal toxins in bottlenose dolphins (Tursiops truncatus) stranding during an unusual mortality event in Texas, USA. Harmful Algae 10, 330–336 (2011).
- Tezanos-Pinto, G. et al. Decline in local abundance of bottlenose dolphins (Tursiops truncatus) in the Bay of Islands, New Zealand. Marine Mammal Science, n/a-n/a, doi:10.1111/mms.12008 (2013).
- Tezanos-Pinto, G., Constantine, R., Mourão, F., Berghan, J. & Scott Baker, C. High calf mortality in bottlenose dolphins in the Bay of Islands, New Zealand–a local unit in decline. Marine Mammal Science 31, 540-559, doi:10.1111/mms.12174 (2015).
- Lusseau, D. L., Slooten, E. & Currey, R. J. C. Unsustainable dolphin-watching tourism in Fjordland, New Zealand. Tourism in Marine Environments 3, 173-178 (2007).
- Christiansen, F. & Lusseau, D. in Whale-watching, sustainable tourism and ecological management. Cambridge University Press, Cambridge, UK (eds J. E. S. Higham, L. Beijder, & R. williams) Ch. 13, 177-192 (Cambridge University Press, 2014).
- Lusseau, D. & Beijder, L. The Long-term Consequences of Short-term Responses to Disturbance Experiences from Whalewatching Impact Assessment. International Journal of Comparative Psychology 20, 228-236 (2007).
- Bejder, L. et al. Decline in relative abundance of bottlenose dolphins exposed to long-term disturbance. Conservation Biology 20, 1791-1798, doi:DOI: 10.1111/j.1523-1739.2006.00540.x (2006).
- Filby, N. E., Stockin, K. A. & Scarpaci, C. Long-term responses of Burrunan dolphins (Tursiops australis) to swim-with dolphin tourism in Port Phillip Bay, Victoria, Australia: A population at risk. Global Ecology and Conservation 2, 62-71, doi:http://dx.doi.org/10.1016/j.ge... (2014).